(Versión Web, junio 2021)
ANTONIO GONZÁLEZ CORDERO
Resumen
Se exponen de forma resumida los resultados de los análisis de fauna correspondiente a los yacimientos de la Edad del Cobre de la provincia de Cáceres, cuando la domesticación se haya plenamente avanzada y las especies cazadas apenas constituyen ya una pequeña parte de los recursos necesarios para la subsistencia. De la lectura de estos datos se extraen interesantes conclusiones acerca de las relaciones del hombre con el medio y los animales, sobre todo cuando al entablar una serie de paralelos con la fauna representada en el arte rupestre esquemático, se observa por primera vez una fragmentación del ecosistema entre lo que podríamos denominar un espacio domesticado y otro asilvestrado, donde los animales asumen un protagonismo distinto, pues ese reflejo especular que constituye la pintura, nos da a entender como estas relaciones fueron más allá del simple hecho nutricional. Por otra parte, la comparación entre las especies consumidas o cazadas y las representaciones pictóricas, ofrecen la posibilidad de un contrapunto argumental a las cronologías ya de por si relativas del Arte Esquemático.
INTRODUCCIÓN
En los últimos veinte años las excavaciones de asentamientos Neolíticos y de Edad del Cobre en la provincia de Cáceres, aparte de alguna excavación de urgencia motivada por las obras en la construcción de nuevas vías de comunicación, se han limitado prácticamente a tres yacimientos, los Barruecos en Malpartida de Cáceres [1] la Atalaya en Torrequemada y el Alto del Lugar en Torreorgaz [2]. Esto condiciona bastante a la hora de abordar el capítulo relativo al papel que desempeñaron los animales en la economía de los primeros pueblos productores de alimentos, no obstante, y pese a lo poco excavado, los avances en este sentido han sido considerables, pues afortunadamente contamos con un caudal de información precedente [3] a partir del cual poder entender un poco mejor como el hombre aprovechó estos recursos y como la domesticación en concreto, se convirtió en uno de los factores más importantes de la transformación social [4].
Nuestra intención aquí, es confrontar los datos disponibles para ofrecer una lectura actualizada y asequible de las relaciones entre el hombre, la naturaleza y los animales, como partes fundamentales de su subsistencia, utilizando para ello, los análisis de los restos presentes en los registros estratigráficos, para finalmente intentar entablar una serie de paralelos con el arte rupestre esquemático, cuyo reflejo especular, nos da a entender como estas relaciones fueron más allá del simple hecho nutricional.
1. ARQUEOZOOLOGÍA DE LOS YACIMIENTOS CALCOLÍTICOS CACEREÑOS.
El marco general en el que se encuadra el análisis faunístico comprende un total de siete yacimientos cacereños, donde se han obtenido muestras arqueozoológicas válidas. Es una escueta compilación de la cual han sido excluidos dos yacimientos tan significativos como el Conejar [5] o la Canaleja, porque sus restos no proceden de ninguno de los niveles que componen su secuencia estratigráfica, sino de un revuelto provocado en el primero de los casos por el aprovechamiento de la arcilla del interior para la construcción de una cerca de tapial, y en el segundo, por las sucesivas interferencias antrópicas en el yacimiento [6].
De la muestra analizada, hoy podemos decir que, para no perder la relación entre horizontes culturales, el grueso de nuestra investigación se concentrará en un momento Pleno-Final de la Edad del Cobre de la fase definida como III-IV del registro local, a título general, caracterizada por la presencia en los niveles de ocupación de una peculiar cerámica denominada de pastillas repujadas, anterior a la arribada de los especímenes campaniformes, aunque convivirá con ellos en la fase final. En tiempo real, las fechas han sido proporcionadas por el Cerro de la Horca 4215+-100 BP cal BC 2916-2459; Cabrerizas 4060+-85 BP cal BC 2700-2490 y Atalaya con dos fechas, la primera de 4080+-35 BP cal BC 2861-2492 y la segunda de 4120+-35 cal BC 2872-2577, nos situaría en un intervalo de tiempo distribuido a lo largo de la segunda mitad del III milenio a.C.
El uso de las informaciones procedentes de este horizonte tiene como objetivo dar cohesión a nuestros datos, pero ello no obsta para que hagamos uso de la información que pueden aportar secuencias más antiguas de otros yacimientos extremeños, cuando, intentemos abordar el origen y el uso que se hace de una determinada especie o comentar los niveles específicos de los contenidos faunísticos de cada yacimiento, pues la interacción resulta primordial para establecer las líneas evolutivas en el manejo de cada una de las especies.
Para comprender, sin embargo, la incidencia de las categorías taxonómicas en el registro final de los poblados de la Edad el Cobre, hemos procurado trabajar las conclusiones contrastando la información ofrecida por los siguientes yacimientos: Cerro de la Horca (Plasenzuela), Cabrerizas (La Cumbre), Castrejón, Castillejos II (Trujillo), Atalaya (Torrequemada), el Alto del Lugar (Torreorgaz) y Los Barruecos (Malpartida de Cáceres) [7]. La intención es, en el escaso espacio disponible, ofrecer desde la perspectiva económica, las consecuencias que tuvo el uso y explotación de los animales en el modo de vida de las poblaciones humanas de aquella época, circunscribiendo el trabajo a un marco geográfico reducido, capaz de facilitar su operatividad. No obstante, en el capítulo dedicado a este fin en nuestra tesis [8], confrontábamos los datos del registro extremeño con otras ochenta y cinco estaciones peninsulares, cuyo registro artefactual, venía a ser cronológicamente coincidente para proporcionar una apoyatura más sólida, a lo que no dejarán de ser conclusiones provisionales y referente de futuros hallazgos.
1.1. Cabra (Capra hircus) y oveja (Ovis aries)
Cabra-oveja, es una pareja de difícil distinción a nivel osteológico, cuya primacia marca la base de la dedicación agropecuaria al constituir el género mejor representado en casi todas las parrillas faunísticas de los yacimientos peninsulares desde el mesolítico. En esas fechas se creía introducido en Europa donde supuestamente no existía el agriotipo de las caracterizó [9], pero el cúmulo de datos recopilados en los últimos años permite suponer que existía una cabra de pequeño tamaño ya desde el paleolítico superior cuya domesticación culminaría en el mesolítico [10]. Esto reforzaría la idea de una mayor antigüedad del componente pastoril sobre el propiamente agrícola, independientemente de su conversión en la cabaña doméstica preferente, como así lo corroboran las estratigrafías neolíticas de la cueva de la Carigüela [11], los Murciélagos, Los Mármoles [12], o La Sarsa [13], donde se observa ya una apreciable distancia entre aquellos y las especies salvajes.
En los Barruecos, el último muestreo realizado a raíz de las excavaciones del 2001 y 2002 en las distintas franjas de ocupación, se ha podido separar la fauna por niveles [14], e igual que en ejemplos anteriores, ofrece ya una reorientación hacia el consumo de ovicápridos con índices variables, pero con un incremento progresivo de su representatividad, traducidas en unos tantos por ciento que oscilan entre el 25% del Neolítico Antiguo, al 50% del Neolítico Medio, 30% del Final y el 23,5% de las fases iniciales del Calcolítico. Esta lectura viene a coincidir con la que ofrecen los demás yacimientos extremeños, cuyo baremo se establecerá en las fases subsiguientes entorno al 21,66%, porcentaje que puede subir aún más de considerar casos particulares de la trascendencia del Cerro de la Horca con un 28,50% en la fase II y un 27,74% en la fase III-IV y como caso intermedio el de la Atalaya con un 26,6%. La media en la Península con respecto a Extremadura es sin embargo más baja, del 13,43%, pero sigue siendo igualmente el número más alto con respecto a todo el espectro arqueofaunístico, que se sitúa siempre en niveles inferiores. No obstante, su identificación en 65 de los 85 yacimientos cotejados a nivel peninsular, habla de la importancia alcanzada, máxime cuando su dispersión cubre algo más de dos tercios del mapa peninsular.
Uno de los problemas que plantea la distinción entre cabra u oveja es el tamaño de las muestras, por lo general recogidas en un estado de fragmentación superior al que experimenta cualquier otro animal tras haberse sometido a un troceado minucioso para su consumo. A pesar de ello, hay unas pautas reconstruibles, por ejemplo, la estructura de la cabaña sacrificada afecta al 40% de entre los 12 y 24 meses, la otra mitad se reserva para aprovechar la lana y la leche [15], mientras a una décima parte, seguramente con vistas a la reproducción, se le da muerte cuando llegan a la edad adulta.
En la geografía ibérica hay lugares donde la incidencia de esta cabaña es mayor, llegando a superar el 46,3% del testado faunístico en Almizaraque [16], el 55% en el Cerro de la Virgen [17] y así en un conjunto de yacimientos normalmente situados en la mitad meridional. En todos esos lugares es posible que la predilección por esta cabaña forme parte de una estrategia adaptativa, quizá en respuesta al continuo proceso de degradación y deforestación que parece desprenderse de los análisis polínicos. Así lo hemos apreciado también en las oscilaciones en la cabaña doméstica de Papa Uvas, especialmente la de ovicápridos, que pasan en la fase I del 32% a 39% en la fase IV [18], aunque la discusión se mantiene en si el grado de alteración del ecosistema es un producto del hombre, o por el contrario se debe a un aumento de la aridez en el sur peninsular.
Hay que recalcar, por último, que la importancia de esta cabaña viene dada por el rápido desarrollo y el ambivalente aprovechamiento que puede hacerse de las mismas, donde si es importante la obtención de carne destinada al consumo, también lo son aquellos productos que se obtienen de ellas, lana, piel, huesos, leche, y los productos derivados de esta última; a ellos cabe añadir otras posibilidades rara vez contempladas, las cuales tienen que ver con la eficaz actuación de esta especie en los espacios amontesados o boscosos, donde su introducción, posibilita el mantenimiento de la pradera castigando al piso arbustivo y la fertilización del terreno como paso previo a la siembra, sin desdeñar tampoco la facilidad y rapidez a las que se presta su desplazamiento y las amplias distancias del territorio que son capaces de cubrir en una jornada. En Extremadura resultaron especialmente útiles, porque entre las especies domésticas, se convirtieron en las mejor adaptadas a los pastos secos, a la vegetación dirilignosa de esta región y a los terrenos de carácter abrupto y montañoso donde se emplazan normalmente los poblados calcolíticos cacereños. Por último, Chapman [19] nos recuerda a propósito de su capacidad reproductiva y a su capacidad de crecimiento entre un 18% y 33% anual, que constituyen la opción más lógica y racional para el mantenimiento de una comunidad.
En cuanto a la discriminación antedicha entre oveja y cabra, la primera identificación clara de oveja tuvo lugar en niveles neolíticos de los Barruecos [20] a través del análisis morfológico de la clavija de los cuernos. Más tarde el mismo elemento anatómico servirá para su nueva detección, esta vez en niveles calcolíticos de la Atalaya de Torrequemada [21]. Esta segregación es interesante, tanto en cuanto se infiere que ya se hacía uso de ambas especies, Ovis aries y Capra hircus, desde épocas muy tempranas, hasta el punto que ocupa un lugar destacado en su explotación por parte del hombre con respecto al resto de los animales, su poca agresividad, tamaño manejable, su carácter gregario, temprana madurez sexual, alta adaptabilidad y elevada tasa reproductiva debieron de influir notablemente en su elección, explicando por otra parte, los altos niveles de representación en los yacimientos, normalmente más de una cuarta parte de los especímenes identificados en los yacimientos cacereños.
1.2. Cerdo doméstico (Sus domesticus) y jabalí (Sus scrofa).
El ganado porcino constituye otro de los pilares de la alimentación de las comunidades prehistóricas, por esta razón se hará presente desde el Epipaleolítico en Nerja [22], manteniendo estable su representatividad en el Calcolítico a nivel general con un 9,28%, cifra que en Extremadura se duplica, propiciada quizá por las excelentes características del medio para la cría de esta cabaña. El techo se alcanza en Cabrerizas con un 26,08%, lo que supone una cuarta parte de toda la actividad ganadera del sitio, muy lejos no obstante de 45,5 que de promedio alcanza en el yacimiento de Mercador en el Alentejo [23].
Igual que ocurre con los ovicápridos, contamos con suficientes pruebas para pensar que esta especie fue seleccionada para su crianza, entre otras razones por la rapidez de crecimiento, superior a la de cualquier animal doméstico, de manera que en un año o año y medio se convertían en animales aptos para el sacrificio con un óptimo aprovechamiento. Por otro lado, su capacidad reproductiva les permitía aportar un gran número de crías y la alimentación omnívora aprovechar desde los infinitos recursos del medio boscoso, a subproductos agrícolas de poco valor o eliminar parte de la basura orgánica que se generaba en una comunidad.
En Extremadura su consumo aún no tiene un patrón claro de explotación de la cabaña, pues la proporción de edades, siguiendo los dos estudios existentes [24], en la mayoría de los yacimientos presenta un equilibrio entre animales adultos e inmaduros, con un grupo de individuos menores de un año y otro ligeramente mayor de adultos cuyo sacrifico tan solo duplicaría la edad de los anteriores, si acaso en la Atalaya, quizá de forma circunstancial hay una mayor abundancia de suidos sacrificados a temprana edad. Tampoco puede deducirse gran cosa de los estudios biométricos, cuyas mediciones bastan simplemente para confirmar que entran dentro del dominio de variación del cerdo coetáneo peninsular.
El material óseo de los suidos aparece muy fragmentado en los yacimientos extremeños, ya que estas especies han sido objeto de un sistemático consumo alimenticio que les situó en el segundo lugar de la fauna doméstica detrás de los ovicaprinos. Queda pendiente aún una reconstrucción morfológica completa de los ejemplares extremeños a través de los cuales se pueda determinar el tamaño y alzada de los mismos.
Entre las piezas recuperadas resulta muy complicado distinguir cuales pertenecerían a su homónimo silvestre y cuáles a la variedad Sus scrofa-domesticus. No obstante, su presencia es notoria en muchos de los yacimientos excavados, siendo los restos de individuos de edad más avanzada uno de los argumentos a valorar cuando se trata de determinar la presencia del espécimen salvaje.
1.3. Vacuno doméstico (Bos taurus) o salvaje (Bos primigenius).
La especie doméstica de bóvido es la tercera de las especies sobre las que descansa la subsistencia de las comunidades calcolíticas extremeñas con un 8.04%, una cantidad muy similar al 11,85% de media en los 60 yacimientos calcolíticos estudiados a nivel peninsular. Algo menor es la correspondiente a la especie salvaje, cuyo porcentaje es de 3,98% en los asentamientos del complejo de Plasenzuela, números igualmente próximos a la media peninsular del 3,75%, si bien el cúmulo de yacimientos donde ha sido aislada esta especie, un total de 19 lugares, arroja una cifra significativamente menor que la anterior, aunque hemos considerado interesante, cuando han podido establecerse las diferencias entre ganado doméstico y uro, presentarlas por separado. En el caso de la Atalaya, donde no se ha podido realizar esta discriminación, su porcentaje es muy elevado, de un 23,5%, la cifra más alta de los yacimientos cacereños, de ahí que en el cómputo general la subfamilia Bovinae, adelante cuantitativamente a la de los Equidae.
Estas cantidades son un síntoma de lo avanzado de la domesticación de esta especie en el territorio cacereño, pero contrastan vivamente con las del yacimiento de Fuente de Cantos, donde la presencia del agriotipo salvaje con un 32,95% no sólo es clara, sino de un dominio apabullante con respecto a la especie doméstica, ausente totalmente en los índices. A nuestro entender, resulta ciertamente chocante esta divergencia, puesto que la diferencia no se manifiesta sólo con respecto a los contenidos de los yacimientos cacereños, sino en relación al resto de las estaciones peninsulares. Castaños [25] lo argumenta como un fenómeno propio de la domesticación asíncrono, no generalizado, y diferente según regiones geográficas, pero conocedores del escaso espacio excavado del donde procedía la muestra, estamos a favor de ampliar el área de reconocimiento, pues este fenómeno de concentración de restos del uro salvaje pudo deberse a una acumulación circunstancial, a todas luces insuficiente, para proclamar que el predominio global del bovino en los histogramas de frecuencias porcentuales de Extremadura, vienen producidos por la importancia cuantitativa del yacimiento badajocense [26]. Otra cuestión bien distinta son las cifras de ganado bovino alcanzados en Valencina o Zambujal, cercanos al 50%, pues se trata de especies domésticas, subrayando, la importancia de la cría de terneros y novillos como forma de especialización en dichos yacimientos [27].
Llama la atención el contingente numérico de animales viejos en Cerro de la Horca, con valores similares al Cerro de la Virgen, Valencina de la Concepción, Jovades y Niuet, un asunto en relación a la probable utilización de los ejemplares para el trabajo [28] o su optimización para el aprovechamiento de la carne. Lamentablemente no se ha podido comprobar aún si hay unas directrices en el manejo de este ganado, ni siquiera si los metatarsos con mayores desgastes corresponden a ejemplares de un sexo determinado, por ejemplo, de machos que insinúan su uso como fuerza de tracción, cuya implicación en relación de fuerzas productivas, puede ser determinante a la hora de valorar también los procesos de transformación agraria.
Una última cuestión es la distribución anatómica de restos, escasa en lo que atañe a elementos del tronco procedente de los uros, posiblemente porque el troceado y despiece fue efectuado fuera de los yacimientos y tras ello acarreadas las partes para su consumo. Pruebas de la existencia de esta especie seguirán obteniéndose en el Orientalizante tardío de Medellín 600-500 a.C. [29]) e incluso en época romana, cuando debió de extinguirse completamente en la Península.
1.4. Caballo doméstico (Equus caballus) y caballo salvaje (Equs ferus).
Fueron necesarios los análisis de fauna de Zambujal o Fornea a cargo de Von den Driesch y Boessneck, para desmontar la teoría de Schüle que negaba la existencia entre la fauna postpleistocéncia de caballos salvajes en la Península hasta la etapa campaniforme, fechas en las que según Harrison [30] se produciría una expansión a gran escala incorporándose como animal doméstico. Años después, los hallazgos de équidos en yacimientos Neolíticos y Calcolíticos se han multiplicado hasta el punto de ser reconocidos en 55 de los 92 test de fauna realizados hasta el momento de escribir estas líneas, pasando a ocupar el tercer lugar en orden de importancia con un 9,48% sobre el total de restos, sin que hoy podamos albergar la menor duda de que muchos de ellos fueron captados de la fauna salvaje como una opción dietética más, pues son frecuentes las huellas de cortes y los astillamientos intencionados en los huesos para aprovechar la carne y la médula, aunque es precisamente la ausencia de estas marcas y la escasa fragmentación de sus huesos en la Atalaya [31], lo que despierta sospechas sobre un uso funcional temprano de la especie.
Esta sospecha sin embargo no basta para hablar claramente de domesticación, pues los criterios morfométricos que generalmente se aplican para conocer el estado de las especies resultan muy poco útiles. Pese a ello, hay quienes sitúan la domesticación del caballo a finales del Neolítico, muy cerca de la frontera del V milenio [32], o más concretamente en la transición Neolítico-Calcolítico [33], de ser así, significaría que en la Península se habría producido la domesticación de forma independiente a Europa Central, y aunque recientemente se ha insistido en la segunda mitad del tercer milenio como el periodo factible para que se hubiera producido la domesticación efectiva [34], se ha de reconocer que no hay total seguridad, hasta la Edad del Bronce, cuando su presencia se puede deducir ya de la presencia de arneses y piezas de bocados.
Las cronologías de los yacimientos extremeños excavados y con fauna, han puesto sobre la mesa este problema, donde los patrones de mortandad, el tipo de aprovechamiento y el número de ellos se han convertido en excelentes argumentos indirectos para demostrar, como con respecto a fases anteriores, donde su presencia fue residual, hubo un sustancial cambio de interés del grupo humano, suficiente para reconocer el principio de un proceso de domesticación [35]. Si este razonamiento se admite, Castaños [36] certifica que el estatus de esta especie en el Cerro de la Horca, Cabrerizas, Castrejón, sería el de especie doméstica, al menos en la forma inicial, aunque persiste la duda de si los altos índices registrados, no responderían a condiciones más favorables para esta especie en la región. Al respecto insiste Castaños [37], las muestras de los yacimientos extremeños con valores superiores al 15,71% tienen un interés primordial, tanto por la cronología, como por su abundancia, lo que les convierte en uno de los yacimientos suroccidentales más destacados en este aspecto, tal vez síntoma premonitorio del papel que habrán de tener en la economía de etapas posteriores, sino lo son ya de iniciales especializaciones ganaderas.
1.5. Perro (Canis familiaris).
Su presencia en yacimientos Calcolíticos extremeños sólo ha sido advertida hasta el momento en Los Barruecos y en la Atalaya, donde con un 0,3 y un 0,7% forma parte del grupo con mínima representación, ligeramente por debajo del 4,74% que ofrecen en conjunto los yacimientos peninsulares. Su aparición es una cuestión ampliamente debatida, por lo que no insistiremos en ello [38], sólo avanzaremos que la presencia de los mismos en determinado contexto, es considerado un síntoma inequívoco de la consolidación de la domesticación.
En los últimos años su presencia ha sido detectada en numerosos yacimientos, muchos de ellos en fosos, silos o sepulcros [39], donde el tratamiento del animal, enterrado entero con bastante frecuencia y a veces junto a restos humanos, se reviste de un especial significado. Ejemplos de este tipo los podemos encontrar en la Gruta de Marizulo en niveles del Neolítico
Inicial y en el sepulcro del Camino del Molino (Murcia), donde los restos de 1300 individuos inhumados, son acompañados por 50 esqueletos completos de perro, datados en la segunda mitad del tercer milenio [40].
Su consumo, aunque hay algunos casos contrastados [41], no fue una práctica corriente, como demuestra el hecho de que en muchos lugares donde fueron recuperados huesos, estos procedían de enterramientos o el estado de los mismos no presentaba huellas de cortes ni abrasiones características de las osamentas cocinadas o despiezadas, hecho sintomático ya de una consideración utilitaria probablemente vinculada a la caza y al pastoreo, tal vez el de cabras y ovejas, pues de los 24 lugares donde hacen acto de presencia estos animales, en 21 se hallan presentes las mismas.
1.6. Ciervo (Cervus elaphus).
La caza mayor persigue básicamente ungulados salvajes, y entre ellos, el ciervo reúne la mayor cantidad de partes, con unas cifras en Extremadura a veces superiores en un 3,93% a la de bóvidos domésticos, y con unas diferencias que podrían ser mayores de no producirse caídas de hasta cinco puntos en el índice porcentual en Los Barruecos, pues incluso entre el complejo de Plasenzuela y Castillejos de Fuente de Cantos los valores se sitúan entre 17,35% y un 17,58%. Otros datos alentadores de la importancia de la caza de este animal son los porcentajes de los referidos asentamientos cacereños, donde nunca bajan del 10,60% y sin embargo se duplica con el 20,68% de las cifras de Castillejos, siendo igualmente altas en la Atalaya, superiores a las de équidos, aunque bastante por debajo de la serie doméstica habitual.
Respecto a la frecuencia de edades de los ejemplares cazados, se constata una predilección por los individuos adultos, cuya distribución ósea, según las distintas partes del esqueleto, es indicativo de que la mayor parte de los animales llegaron enteros al asentamiento [42]. El aprovechamiento es máximo, y si se prefieren los machos adultos, es seguramente porque la cornamenta constituye una excelente materia prima para la fabricación de herramientas.
Hay punzones y mangos fabricados en este material, huellas de trabajo en una ulna y cuernas de desmogue con abrasiones preparatorias, pero no se menosprecian otras partes del esqueleto tales como las falanges y tarsos empleados en el Cerro de la Horca para fabricar idolillos, una elección de manifiesta intencionalidad, seguramente relacionada con las virtudes adjudicadas a este animal.
En algunos lugares como la cueva del Nacimiento, se ha hablado de la semi domesticación de esta especie [43], intentando relacionarlo con el caso del reno del norte y los lapones, pero nada parece indicar que en Extremadura sucediera algo parecido, la disminución de tamaño observada en las especies más occidentales de la Península parece ajustada a un gradiente geográfico y climácico, razón por la cual los ciervos de Extremadura tiene un parecido mayor con los de Zambujal que con los de Levante español [44].
1.7. Lepóridos: conejo (Orictolagus cunniculus) y liebre (Lepus capensis).
Uno de los problemas planteados por esta especie tiene que ver con sus costumbres fosoras, es decir la excavación de madrigueras que pueden haber aportado sin el intermediario humano, una porción de sus restos a los niveles del asentamiento, bien porque se produjo su muerte en el interior de las mismas, o porque fueron consumidos por otros animales como el zorro dentro de su cubil. Pese a este imponderable, la repetida presencia de los mismos en 37 de los 85 yacimientos del análisis peninsular, les garantiza un lugar destacado dentro del abanico de las especies consumidas, aproximándose por primera vez en los taxones extremeños con un 7,44%, a la media del registro peninsular donde alcanzan el 7,37%. Nadie ha aventurado hasta el momento una hipótesis de interceptación del conejo como especie domesticada en los niveles de la Edad del Cobre, porque para eso se necesitaría la recuperación de cráneos en buen estado, al residir en estos las principales diferencias entre ejemplares salvajes y domésticos, una circunstancia poco habitual entre el material arqueozoológico de esta especie, propensa por su liviandad a una pérdida acelerada, si las condiciones del terreno donde se conserva no son las adecuadas. Por esta razón, a cualquier dato relacionado con ellos hay que contemplarlo con unos estrictos márgenes de reserva, por cuanto queda por determinar el porcentaje procedente de la caza, de su domesticación, o si su presencia es ajena a la intervención humana.
En los yacimientos controlados de Extremadura, el conejo presenta una irregular intercepción de ejemplares, con las variaciones de consumo más irregulares entre asentamientos pasando de 2% a un 8% entre las dos Fases calcolíticas del Cerro de la Horca, una varianza entre especies achacable seguramente a las malas condiciones de conservación y al deterioro que sufre una osteología tan frágil en los niveles más antiguos y que pueden asimismo desvirtuar el resultado final sobre el consumo de estos animales.
En los Barruecos se instala con un 15,3% en el tercer lugar de la parrilla de especies que forman el repertorio dietético de los pobladores del sitio, el mayor registro de estas características en la región, sólo comparable con Cerro de la Virgen y Zambujal, donde la importancia que tuvo parece realmente exagerada.
El conejo sigue teniendo una gran importancia económica en la Península Ibérica y en la propia Extremadura, donde a pesar de la recesión del último siglo provocada por enfermedades, continúa siendo abundante, incluida el área de Plasenzuela, donde genera una riqueza anual importante en el capítulo de actividades relacionadas con la caza.
La liebre y en conjunto de animales subsiguientes forman un grupo cuyos índices de representación rara vez superan el 1% en los yacimientos; se limitan por tanto a apariciones mínimas cuya obtención se debe seguramente a una caza de corte oportunista, dando lugar a minúsculos restos susceptibles de ser computados. En Extremadura se hallan en torno al 0,79%, por debajo valor medio peninsular que alcanza el 2,96%. Debe no obstante ser un animal con un nicho de actividad muy amplio, pues en todas las zonas del muestreo siempre se encuentra presente algún espécimen.
1.8. Corzo (Capreolus capreolus).
Es un importante indicador de formas paisajísticas, al ser el usuario característico de los montes abiertos. Fue menos estimado que el venado, quizá debido a su corto número en comparación con otras especies y a la natural dificultad de su rececho. Prefiere los bosques caducifolios, pero se ha adaptado bien a las masas de la foresta impenetrable del monte mediterráneo, aunque en Extremadura campa muy lejos de los territorios que en el III milenio formaron parte del área natural de su distribución.
Aparece en casi todos los yacimientos extremeños, pero su número nunca ha llegado a superar 1% del total de las especies consumidas, es por tanto una caza ocasional, sorprendente por el bajo interés que despierta, cuando comparada al venado su carne es de similar o superior calidad, si bien sus defensas son menos aprovechables debido al reducido tamaño.
1.9. Cabra montés (Capra Pyrenaica).
Esta especie, confinada actualmente en nuestra región a las sierras del Sistema Central, en el intervalo de tiempo que aquí contemplamos, debió de proliferar por espacios más amplios y diversos, pues sin abundar excesivamente, se hace presente en la mayor parte de los yacimientos analizados, desde Fuente de Cantos a Los Barruecos. El material de examen es no obstante bastante escaso, quizá porque al igual que sucede con el corzo, su hábitat se ciñe a reductos de gran aspereza, sobre todo si se ha de trasportar la carne, o sencillamente porque al no tener la calidad de la especie doméstica y necesitar de un rececho complicado para su captura, la hace menos apetecible. El índice con el que se presenta en Extremadura es de 0,28% y en la Península de 1,77%, si bien en relación a estos últimos, los yacimientos donde se menciona a esta especie constituyen un 22,22%.
1.10. Lince (Lynx pardinus).
Ha sido detectado en 12 yacimientos de la Península, emparejando su representatividad a la del zorro con un valor de 0,9% deducible de su presencia en Fuente de Cantos y la Atalaya. Debió de ser cazado por las mismas razones que el anterior, quizá con más asiduidad, al tratarse de un depredador habitual de otras especies cazadas por el hombre.
1.11. Zorro (Vulpes vulpes).
Sin interés culinario, el zorro fue cazado para obtener su piel y como medio de recortar la competencia en un territorio por otras especies codiciadas para el hombre. En Extremadura está presente en la fase III-IV del Cerro de la Horca y en Castillejos I de Fuente de Cantos.
Alcanza un 0,045% de la representatividad total, en línea con los valores típicos de esta especie en la Península.
1.12. Tejón (Meles meles).
Se encuentra en el Cerro de la Horca y Barruecos con valores del 0,20%, cantidad meramente testimonial. Pesa sobre ellos la sospecha de que algunos de los restos procedan de una deposición por muerte en el interior de la tejoneras o cados. En el resto del país los diez yacimientos donde figura, reducen su representación peninsular al 1,77%.
Este animal, igual que el zorro, el lince, pudieron ser cazados con más frecuencia del expresado en los índices, pero a diferencia de la caza mayor, acarreada hasta el poblado porque interesaba tanto su consumo como la obtención de partes anatómicas para ulteriores elaboraciones, los cuatro últimos, sólo debieron de interesar al cazador principalmente por su piel, tendones o dientes, consecuentemente el despiece y abandono de los restos debía de realizarse en el mismo lugar donde se cobraba la pieza, impidiendo su incorporación de otras partes del esqueleto a los basureros de un asentamiento, de donde habitualmente procede nuestra provisión de huesos predisponiendo a una lectura un tanto sesgada del comportamiento real de consumo de estas especies.
1.13. Otros
Entre los mamíferos podría citarse además al Gato salvaje (Felix silvestris) como resultado de una captación en Fuente de Cantos. Con un 0,02% es una de las especies menos representada en el cómputo cinegético, tan solo por encima del lobo y del oso, ausentes de nuestros recuentos, pese a ser una fauna coetánea.
El resto de la fauna empleada para su consumo tuvo origen también en la pesca y recolección de moluscos de río. Un artículo sobre este tipo de fauna en yacimientos del Campo Arañuelo [45] confirma las noticias que ya tenían acerca del hallazgo de algunos especímenes en el Cerro de la Horca y el Conejar, ampliando en una docena la lista de yacimientos donde se habían conservado las valvas de moluscos recolectados, principalmente de las especies: Unio crassus, Margaritifera auricularia, Potomida littoralis y Unio pictorum, también conocidos como náyades o mejillones de río, cosechados en los fondeaderos de ríos cercanos y trasladados a los asentamientos para ser consumidos, aunque el impacto en la alimentación no debió pasar de un nivel de complementariedad.
La presencia de anzuelos, pesas de redes se convertían por otro lado en firmes testimonios de la pesca de especies, cuya fragilidad osteológica había esquivado su detección [46]. Tan sólo una vértebra de anguila con huellas de fuego en la Canaleja constituía el único indicio real del uso de la pesca como complemento dietético para aquellas poblaciones.
No existen de momento otras pruebas que nos hablen del consumo de roedores, batracios, ofidios u otras especies, y sólo entre las aves tenemos dos testimonios que pueden remitirnos a usos muy diferenciados. En cuanto al primero, un pequeño resto en la Atalaya [47] de un único espécimen de ave no identificada, con huellas de mordeduras probablemente humanas, sea quizá el único indicio de una captación dietética más amplificada de los restos hasta ahora obtenidos nos permiten ver. El segundo quizá tenga que ver, dentro de un orden funcional, con uso de algunos huesos, en concreto el cúbito de un águila, único de los fragmentos de ave recuperado en niveles de la Fase III-IV del Cerro de la Horca. Las características morfológicas y tal vez simbólicas de los huesos de las alas de esta especie, de extraordinaria longitud, rectilíneos, huecos y de sección circular, les han convertido en la materia prima ideal para construir instrumentos de viento a modo de siringas o flautas de pan, con ejemplos tan ilustrativos como los de la Cova de L´Or, un yacimiento del Neolítico Antiguo [48]. El fragmento del Cerro de la Horca también es hueco, pero dado estado de fragmentación resulta muy difícil afirmar que tuviera un empleo similar al del yacimiento de Beniarrés o fuera empleado como un pequeño recipiente para algún tipo de substancia, pues de lo que si estamos seguros es que el intenso patinado que presenta no se corresponde con un simple resto de fauna consumida.
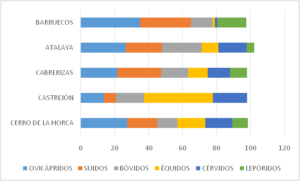
Cuadro nº 1: Composición y porcentajes de los grupos taxonómicos más frecuentes representados en los yacimientos calcolíticos cacereños.
- FAUNA DOMÉSTICA.
En uniforme preliminar referido a la fauna prehistórica extremeña Castaños,[49] señalaba dentro del conjunto de especies identificadas en la región, tres bloques de distinto significado arqueozoológico que nosotros hemos compartimentado en seis dependiendo de la importancia desempeñada por cada uno de ellos.
Por un lado, los animales domésticos o con visos de domesticación: oveja-cabra, cerdo, caballo, vaca; en segundo término los ungulados salvajes objeto de caza: ciervo, uro, jabalí, cabra montés, corzo; y un tercer bloque de contenido diverso caracterizado por cuatro subgrupos: el de lagomorfos (conejo y liebre) aún con porcentajes importantes en la parrilla de yacimientos; el de los carnívoros: tejón, lince, zorro, gato montés, perro, con representación por debajo del 1%; el de las aves y por último el de moluscos con un porcentaje ligeramente superior a últimos. En total 16 referencias a macromamíferos, siendo muy raras las referencias a la avifauna y pequeños vertebrados, de los que apenas existe información.
La suma de los distintos bloques, nos depara unas cifras que son de gran interés para comprender cuales fueron las preferencias dietéticas de aquellos pueblos, orientada en el caso extremeño hacia el consumo de animales domésticos. La banda donde quedan expresados dichos niveles es bastante elocuente, pues todas aparecen instaladas en torno a valores similares, entre los 68,97% de mínimo en Castrejón y el 81,1 de máximo en la Atalaya, con una media porcentual del 75,03%, es decir que casi tres cuartas partes de la aportación cárnica procedía de recursos de fauna controlados. Sorprende la escasa variación existente entre los siete casos analizados, lo que en cierto modo no sólo corrobora la hipótesis de que estamos ante grupos humanos con una economía preferentemente ganadera como sucede en otras áreas peninsulares, sino que existe un comportamiento subyacente de explotación y consumo bastante similar entre todos ellos.
En el examen de las distintas cabañas, ha sido posible identificar además, las especies de cría preferenciales, con una pauta claramente dominante en favor de los ovicápridos con un 25,82%; le siguen en importancia la cabaña porcina que con un 21,16% absorbe casi otra cuarta parte de la producción pecuaria; la bovina con 16,61% y por último la caballar con un 13,33%. Esta dedicación supuestamente tan lineal, lleva aparejada sin embargo ligeras fluctuaciones que pueden encauzarnos hacia la comprensión de formulaciones de organización económica mucho más complejas, como las que pueden deducirse del examen de las áreas de poblamiento sincrónico y relativamente próximo a Plasenzuela.
En dicha zona y en la cercana Torrequemada, los sitios arqueológicos, por ejemplo, coinciden con los resultados enunciados hasta ahora a nivel general, pero a título particular manifiesta una especial preferencia por alguna de las cuatro cabañas domésticas: el Cerro de la Horca y Atalaya ovicápridos, con un índice para la fase III-IV entre el 27,74 % y el 26,6 %, de hasta 14 puntos por encima de los restantes; Castrejón ganado caballar, con un índice del 41,37% superior a cualquiera de las frecuencias extremeñas y una de las más altas de la Península, y por último Cabrerizas, que se aúpa con un 26,37% al primer puesto entre cifras de consumo de ganado porcino. ¿Podría considerarse un atisbo de especialización territorial?, posiblemente, pero en las circunstancias en las que se desarrolla la economía de subsistencia, obligó a cada una de ellas a no descuidar la crianza de las restantes cabañas, pues estas contribuyen a diversificar los recursos y por tanto contribuyen a reforzar la seguridad alimentaria.
En relación a la composición porcentual, las preferencias guardan también un orden lógico en relación con la capacidad de mantenimiento de las cabañas, pues dependen del tipo y cantidad de vegetación, de las prácticas agrícolas y en líneas generales de la actuación del hombre sobre el medio, es decir que la situación de las mismas variará en función de las exigencias de cada una de ellas y el espacio donde interactúan. No es lo mismo, por ejemplo, mantener una oveja o una cabra que un caballo, la cantidad de forraje que demanda el tercero limita la cría de estos últimos a gran escala, dejando mayor margen a las primeras que son menos exigentes. Esto como se ha adelantado, simplifica la explicación buscada en varios yacimientos, donde se produce fuerte incremento de ovicápridos, cuando por distintas causas se asiste a un incremento de la aridez. Hay pues una razón de complementariedad y de estrecha dependencia entre el comportamiento del ganado y el medio, cuya relación será determinante en la configuración de ambos.
La distribución de la que hemos hablado, sólo está sujeta a una modificación en términos arqueofaunísticos cuando se tiene en cuanta el peso de los huesos, o su traducción en volumen de carne, es entonces cuando se producen modificaciones notables y el caballo o el ganado doméstico bovino se aúpan por encima de ovicápridos y suidos.
A un nivel más general, el orden en el que se manifiestan las cabañas repite su actuación en innumerables asentamientos, entablando notables paralelismos el Cerro de la Horca por ejemplo con Monte da Tumba, especialmente la fase Ib, con un predominio de la oveja sobre el cerdo [50], La Peña de los Gitanos de Montefrío, Valencina de la Concepción o Zambujal, todos pertenecientes a la geografía suroccidental, donde se pone de relieve de nuevo la polarización ganadera en torno a cuatro o cinco agriotipos, ovicápridos, suidos, bóvidos y équidos, por delante del ciervo que sigue ocupando una posición de ventaja con respecto al resto de las especies salvajes, a excepción de los leporidae. Frente a este estilo de producción, se encuentra el de las regiones más áridas del sureste, donde, el predominio de los ovicápridos con respecto a otras cabañas y más aún de la fauna salvaje, es abrumador; pese a ello se encuentran parecidos sorprendentes, por ejemplo, con Terrera Ventura [51].
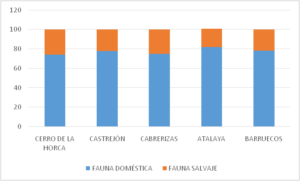
Cuadro nº 2: Histograma con valores comparativos entre la fauna doméstica y salvaje de los yacimientos calcolíticos de la provincia de Cáceres.
3. FAUNA SALVAJE.
En un segundo plano queda la caza, convertida en una aportación complementaria con solo una cuarta parte de la carne consumida, perdiendo paulatinamente importancia a medida que se consolidan las prácticas ganaderas, reflejándose esta tendencia en los cuadros de especies clasificadas en el yacimiento de la Edad del Bronce del Cerro del Castillo en Alange [52].
La nota más característica del recuento efectuado en los yacimientos extremeños, es el elevado índice registrado en el consumo de carne de ciervo, superior incluso al de yacimientos coetáneos del mediodía peninsular y al de alguna especie doméstica como ya se ha comentado. Esta abundancia bastante generalizada en la región, no debe valorarse meramente como una peculiar apetencia gastronómica, sino como la consecuencia de una práctica ecológica y ritual heredada de etapas anteriores. En este sentido, hemos destacado el papel atribuido al animal, cuya predilección no sólo queda patente en el registro arqueozoológico de los yacimientos, sino en las manifestaciones artísticas esquemáticas, donde asumen un claro protagonismo de signo toténico.
La penuria con respecto al anterior de corzo y jabalí, hace que su aportación sea prácticamente anecdótica, tan sólo el uro y su considerable reserva de carne, tentó a los cazadores lo suficiente como para obviar el peligro de su captura, si es que no fueron la consecuencia de batidas selectivas destinadas a proteger los campos de cultivo [53].
Descartamos la idea generalizada acerca de la cual las culturas pastoriles no demuestran inclinación por la caza, cuando prescindiendo de los índices de algunas especies antes mencionadas, esta actividad se puede percibir gracias a la presencia de puntas de flechas, industria que en los yacimientos excavados aventajan o igualan a otras manufacturas del ramo.
La presencia de conejos y liebres siempre resulta problemática, pues pueden tener origen en procesos específicos de tafocenosis o depredación, pero la larga lista de yacimientos y las condiciones de conservación, huellas, roturas y otros signos de manipulación nos han convencido de que una buena parte de los restos son el resultado de capturas realizadas por el hombre. Los índices no otorgan un carácter preferente a los mismos, y mucho menos a la liebre, exiguamente representada en cualquiera de los registros, pero ello no obsta para acreditar el consumo de esta carne como una opción dietética complementaria y muy bien ponderada en épocas de escasez.
De caza selectiva se pueden calificar las capturas de animales que conforman el bloque compuesto principalmente por carnívoros, pues estos son eventuales competidores del cazador y potenciales enemigos del ganado cuyo control fue necesario planificar, procediendo incluso a un exterminio siempre difícil de computar, especialmente en las áreas de pastoreo. La ausencia de restos de un depredador como el lobo en la gran mayoría de los registros de fauna extremeños, quizá hay que tenerla en cuenta en este sentido. Altos índices, de piezas cazadas en una primera fase de Monte da Tumba, puestas en relación con la instalación del poblado [54], pueden ser la prueba más evidente de la realización de campañas de descaste previa a la ocupación de un territorio
4. LA FAUNA EN EL ESPEJO DEL ARTE RUPESTRE.
Es evidente que en el arte rupestre esquemático cacereño existe un conjunto faunístico cualitativamente variado, donde aún resulta muy difícil decidir cuáles son las especies dominantes de ese cortejo, pues más de tres cuartas parte del mismo han reducido la imagen animal a un símbolo donde la especie es indistinguible. Sin embargo, en la parte restante, una observación detallada de todos los conjuntos conocidos, nos lleva a señalar la existencia de un proceso de selección de una serie de animales que podemos emparejar con casi todas las especies identificadas en los yacimientos neolíticos y calcolíticos excavados, aportando más información acerca de la interacción con los seres humanos, sobre todo porque su contenido social y simbólico nos muestra un camino que va más allá de las actividades cotidianas de sus autores.
–Ovicápridos. Prescindiendo de aquellos zoomorfos por cuya presencia cuernos podríamos atribuir vagamente a alguna otra especie, su retrato aparece en bastantes ocasiones, aunque no son pocas las que arrojan sombras de duda acerca de su reconocimiento. Para tratar de superar este obstáculo, una investigación a cargo de García Arranz [55], planteó una fórmula muy sencilla, basada en la identificación como escenas de pastoreo a aquellas donde la asociación entre hombre-cuadrúpedo era evidente y algún atributo o actitud, pudiera revelar la especie del animal representada como perteneciente a una de los animales característicos de la serie doméstica. Así clasificó a las figuras de los paneles I y II del abrigo de Rosa, donde entre dos antropomorfos y un ramiforme se sitúa una pequeña figura con una aparente cornamenta que no duda en identificar como un cáprido. Como escena de pastoreo interpreta otras en el Cancho de la Burra y si en esa ocasión no arriesga a identificar los animales [56] es por la falta de definición de algunos y porque quizá en una investigación previa, el de mejor fijación, aparecía consignado como un cánido [57]. Nadie repara, y con gran nivel de detalle se aprecia en el calco de J. J. García, en dos apéndices que indefectiblemente distingue a las cabras de otra especie, y que no es otro que dos repliegues de la piel en forma de tubo colgando del cuello denominados en Extremadura mamellas, añadidos con mucha precisión al dibujo del animal. Se trata por tanto y con casi total seguridad de una escena de manifiesta relación con el pastoreo, casi idéntica a otra del Paso de Pablo, donde otro animal sobrepuesto a una figura antropomorfa, ha sido retratado con una cornamenta muy corta, explicitando de nuevo la especie caprina con el añadido de las mamellas colgando del cuello, si es que en este caso no se trata de las barbas de un macho cabrío (Fig. 1)
–Équidos. Tal vez nunca hallamos dudado en interpretar como équidos las figuras de animales emparejadas que aparecen en las estelas de guerrero de la Edad del Bronce porque su asociación a un carro permite la elipsis de otros rasgos identificativos del animal. Esto por un lado pone de relieve como el lenguaje de lo esquemático seguía aún vigente al final de la Edad del Bronce y por otro, plantea un problema a la hora de reconocer este animal, pues si
su imagen en la pintura rupestre se limita a ese nivel de sencillez, en esas condiciones, por poner un ejemplo, determinados grupos de animales representados en varios abrigos extremeños junto a un antropomorfo y con trazo similar, pudieran ser interpretados como tales.
Indudablemente una escena de equitación, es decir un caballo con el jinete montado, sería mucho más creíble, de ahí que por este medio se intentara demostrar en una ocasión la representación de esta especie, a partir de tres motivos localizados en la comarca de las Villuercas, no sin antes advertir por parte de su investigador [58] que la fuerte esquematización les convertía en casos hipotéticos. Los dos primeros están encajados en reducidísimos paneles de la cueva del Cancho de las Sábanas y el tercero en el profuso panel del Cancho del Reloj, repitiendo un esquema similar de cuadrúpedo con un motivo en T superpuesto. Como escena de domesticación de un équido se interpreta un panel en Sierra de la Corchuela de Monfragüe [59] y otra en Los Barruecos [60], pero igual que en los anteriores, la fuerte abstracción de las figuras en nuestra opinión dificulta la atribución inicial.
Hubo que esperar al descubrimiento del abrigo Florencio en Retamosa [61], para poder especular sobre la presencia de caballos en la pintura, cuando el filtrado de una imagen del mencionado abrigo reveló, la presencia de una figura tratada con ciertas dosis de naturalismo, donde se muestra parte de la grupa, cola, corvejón y muslos de los cuartos traseros de un animal, desgraciadamente la imprimación posterior de gruesas digitaciones, anula prácticamente la posible visión de la parte anterior del tronco y apenas se distingue una sombra dentro de un borrón de color rojo purpureo (Fig.2).
–Bóvidos. La limpieza de un panel de Monfragüe despertó la posibilidad de que una de las figuras, otrora considerada un cérvido o cáprido por su llamativa cornamenta curvada, pudiera tratarse en realidad de un bóvido [62] y quizá una más en el subgrupo V2 del abrigo Florencio [63], pero la instantánea de mayor valor, pues nos informa además de la posibilidad de domesticación de la especie, es la representación de un bóvido en el Paso de Pablo [64], donde aparentemente, un individuo, tal vez una mujer con faldellín y un tocado en la cabeza, sujeta al animal con una cuerda (Fig. 3). Si descartamos por imposible una escena de caza a lazo de un ejemplar de estas características, obviamente se trata de un control del animal por parte del hombre que lo guía, retratando una de las escenas con mayor carga informativa del mapa pictórico de las Villuercas, pues puede llevar implícito el correlato mítico de la domesticación a cargo de una entidad femenina, asumiendo un papel reiterado ya en la pintura levantina [65].
Acerca de la misma, resulta difícil no entablar estrechos paralelos con otra similar descrita por Jordá, aunque las pinturas del abrigo de la Peña del Escrito del Villar del Humo en Cuenca se engloban dentro del Estilo Levantino. En ella un antropomorfo igualmente femenino lleva cogido de una cuerda un ternero que a modo de simulacro se presenta como una ofrenda ante un supuesto dios toro [66]. Otros antropomorfos sujetando por el cuello o el hocico con una cuerda a animales, han sido halladas en el abrigo del Tío Campano [67] y Selva Pascuala, y aunque en ambos casos se trata de caballos, se llegó a la conclusión de que en el primero se trataba de una caza a lazo, y en el segundo, contradiciendo lo que en su día expuso Beltrán [68], de una escena de domesticación, pues la rienda acaba en el hocico del animal [69]. Más claro parecen las reproducciones de El Canjorro de Peñarrubia, cuyo calco ofrecido por Breuil [70] no muestra una figura en esta actitud, sino varias, donde el dominio del hombre sobre el animal va implícito en la docilidad del rocín.
Caza o domesticación en muchas de las pinturas de este tipo, son un dilema aún no resuelto, ni siquiera con la ayuda de los métodos analíticos de estilo-temática que proponen una ruptura total de este tipo de estampas, por otra parte, clásicas del Estilo Levantino y del Esquemático, asignándoles simplemente el calificativo de seminaturalistas o postpaleoliticos [71].
Guardarían, eso sí, cierta concordancia con el sustrato de carácter narrativo, pero en esencia, su encaje pretende ser un paréntesis entre una y otra fase, una especie de cajón de sastre, pues ni el estilo, ni lo que se narra, concuerdan con las respuestas que la arqueología por el momento ha sido capaz de ofrecer, con respecto al tema de la domesticación de bóvidos y équidos en la Península Ibérica. Por esa razón cuando se habla de cronologías para estas pinturas, la frontera de IV milenio a. C., en fechas sin calibrar, parece aún la más adecuada.
–Suidos: Sólo existe una pintura en los Ibores descubierta hace pocos años [72] y de factura Epipaleolítica en varios de sus detalles, donde un cazador acosa a un jabalí cuya morfología no entraña ninguna duda. Encontrar sin embargo en la pintura esquemática, imágenes de estos animales, es un desafío a la imaginación, pues no parece haber un estereotipo claro que los identifique. La única imagen aproximada, parte más de una conjetura que de una lectura indiscutible y nos remite a un motivo en el Cancho del Reloj acompañado por otro cuadrúpedo, donde dos trazos paralelos sobre el lomo del primer animal sugieren una escena de caza del mismo acosado por un cánido [73].
–Cérvidos: El ciervo ocupa un lugar prominente en las sociedades prehistóricas, respondiendo a una tradición heredada del mundo postwürmiense cuando este animal llego a constituir gran parte de la dieta de muchos pueblos. Por esta razón, alrededor del animal se llegó a construir una mitología cuya expresión más directa la encontramos en el Arte Rupestre esquemático, donde la asociación de imágenes hombre-animal en estereotipadas escenas venatorias, recuerda constantemente una afinidad que trasciende normalmente de hecho económico que comporta la caza y nos introduce en aspectos rituales e incluso iniciáticos de la misma. Es con todo el animal salvaje más representado de repertorio figurativo extremeño, y superior en cuanto a los hallazgos de sus restos en asentamientos a alguna de las especies domésticas, por lo que no debe extrañar la magnitud de sus cifras porcentuales. Independientemente de los valores mencionados, la caza del ciervo y otras especies afines como la del corzo, pudo partir también de la necesidad de aligerar la presión que estos podían ejercer sobre las superficies cultivadas.
En la pintura esquemática es el animal más fácil de distinguir, sobre todos los ciervos machos, frecuentemente representados con la cuerna ramificada con sus adornos de puntas, candiles y corona. Escenas del abrigo II de la Sierra de la Madrastra en Cañamero con una armada de antropomorfos batiendo una manada de ciervos gobernada por un gran macho junto a hembras y varetos es el gran objetivo de los cazadores, expresando con todo rigor las circunstancias que suelen envolver el lance cinegético tantas veces retratado y a veces insinuado simplemente con la presencia de arqueros. Así lo vimos y así lo han percibido otros investigadores [74], con una lectura de mayor detalle solapada en las actitudes de una narración pocas veces repetida en la pintura esquemática de la zona.
Los ciervos son también las representaciones más antiguas de las Villuercas y prescindiendo del grabado paleolítico de la cueva de la Mina [75], por epipaleolíticas dimos la figura de un gran cérvido de carácter naturalista dibujada en color negro e infrapuesto a los motivos esquemáticos del Paso de Pablo [76] y de la misma época, otro gran cérvido en rojo, detectado por primera vez en el calco de Mª C. Rivero [77] oculta tras el panel principal de la cueva del Castillo de Monfragüe, compuesto por una jerarquía de figuras antropomorfas y otro par de figuras de incorporación más tardía, reconocibles por las ramificaciones ahorquilladas de su cornamenta. (Fig. 4)
En Galicia, donde la representación de cérvidos en el arte rupestre es cuantitativamente muy superior a la de cualquier otra especie, son vistos como animales de un fuerte componente simbólico asociado, igual que en otros grupos del arte europeo, a la fecundidad, como animal psicopompo al mundo funerario, e incluso al culto al sol [78]. Esta traducción, era hasta hace poco difícilmente demostrable en nuestra región, hasta la aparición en las Hurdes de paneles e incluso cerámica con imágenes de cérvidos asociadas a imágenes antropomorfas e idoliformes en un contexto mucho más antiguo que el gallego, pues se enmarca dentro de las producciones calcolíticas. Estos descubrimientos darían pábulo a la hipótesis de que el ciervo habría comenzado a tener alguna connotación religiosa, cobrando sentido así otra figura situada bajo un soliforme en el Cancho del Reloj, cuya extensión de la cornamenta se convierte en parte del halo solar [79].
En nuestra opinión, no es tampoco una coincidencia que sean cápridos y cérvidos los animales más representativos, tanto de la fauna doméstica como de la salvaje, cuando los mismos arrastran en sus respectivas parrillas los mayores porcentajes de fauna consumida.
–Corzos. Quizá la figura más comprometida con la imagen de un corzo del álbum cacereño se encuentra en representada en el marco pétreo aislado, por encima de una repisa natural en el Cancho de la Burra, donde tuvimos oportunidad de fotografiarla en año 1985. Se trata de un macho con una cuerna mucho más sencilla y corta que la de los ciervos, propias de un ejemplar maduro, con astas en paralelo y tres puntas bien delineadas, visibles con más detalle en la cuerna derecha, además de un cuerpo más fuerte, compacto y ancho en los cuartos delanteros y una cola extremadamente corta de apenas unos centímetros.
–Cabras montesas. Su caza debió de formar parte del recuerdo de un tiempo cuando junto al ciervo, con un 39% y un 37% respectivamente, sostenían tres cuartas partes de la representación de animales Estilo Levantino [80]. Ahora esa tendencia se ha invertido, y su índice de frecuentación en los recuentos de los asentamientos es puramente anecdótico, siendo así reflejado en los lienzos pétreos. (Fig. 5)
Indubitablemente distinto y diametralmente diferenciado de las especies domésticas por la longitud y curvatura de su cornamenta, su mejor lectura la tenemos en el abrigo Florencio de Retamosa [81]. El retrato de dos cabras una de perfil y otra en perspectiva torcida, nos ilustran acerca de la forma y el tamaño de sus defensas, donde casi se adivinan los medrones de la cuerna. Otro cáprido con la cola recta y la cornamenta doblada se adivina en otro panel de la misma covacha dibujados con notable minuciosidad. Por último, fuera de este abrigo, en la cueva Bermeja de Serrejón, junto a una figura heliomorfa, creemos ver en un zoomorfo con una gran cuerna bifurcada la imagen de otro de estos animales [82].
–Cánidos: la diferencia entre lobo y perro es compleja, aun cuando se produzca una interacción con especies reconocibles como domésticas. La distinción, por ejemplo, entre quién guarda un rebaño o lo ataca, dependerá del interés y la habilidad del artista por reflejar la escena y eso es precisamente lo que nos encontramos en la parte inferior del Paso de Pablo. Se trata de la acción desarrollada en la parte más visible de la mitad inferior del panel, donde se observa una pareja de cánidos con las orejas rectas y el rabo estirado hacia arriba, en una actitud propia de esta especie cuando se muestra activa y confiada. Un tercer cánido precede a una cabra, la cual es reconocible por los cuernos y un rabo muy corto. La ambigüedad de la escena no permite afirmar con rotundidad que se trate de la salvaguarda de un rebaño si se trata de perros, o de un intento apartar del rebaño a un animal si se trata de lobos, pues depende exclusivamente de si el autor de la reproducción de la escena, tuvo realmente en cuenta la etología animal. De haber sido así, para entenderla como un ataque por parte de los segundos, la fidelidad a la acción hubiera requerido representar a la cola de los cánidos recta y horizontal, no curvada hacia arriba como es el caso. Es por tanto una escena subjetiva, pues depende de un único detalle, de cuya formalidad no sabemos si se prescindiría intencionadamente, en cualquier caso, nos sirve para ilustrar la dificultad que ofrecen este tipo de escenas, pues la fidelidad de los detalles hará variar la interpretación de una escena de pastoreo a una de acometida. (Fig. 6)
La presencia del hombre en escenas con cánidos, a veces puede marcar la diferencia y ejercer de árbitro de una discriminación entre perro o lobo. Hay menos dudas cuando se trata de una escena de caza, donde el perro, por ejemplo, presta una inestimable ayuda en la persecución de las presas salvajes. Éste podría ser el caso de Madrastra II y el de otros muchos lugares fuera de nuestra geografía [83], pero el nivel de concreción artística o desarrollo pictórico prestan siempre dificultad a su reconocimiento y es por lo general el contexto el que asegura su discriminación.
Nos queda la duda de si dos figuras pintadas en el Risquillo de Paulino [84] son un lobo y su lobezno, dicho así por la diferencia de tamaño, o son un zorro y su cría. El trazo es muy esquemático, pero permite diferenciar del tronco unas largas patas, una espesa cola vertical estirada, un hocico alargado y unas orejas grandes y puntiagudas. Como otro posible cánido con la cola curvada hacia adelante se interpreta una figura de Cueva Chiquita [85].
–Félidos. El único lugar donde hemos logrado identificar un felino es en la cueva de los Horcones de la sierra de Navezuelas con un nivel de detalle extraordinario [86]. Es la representación de un animal robusto, con el tronco alargado, rematado por una cola muy larga curvada hacia adelante. La cabeza grande con respecto al cuerpo y redondeada, orejas pequeñas, triangulares y hasta un par de vibrisas parecen asomar de su hocico largas, gruesas y caídas. Como motivo pictórico, los gatos no son una de las especies predilectas, pero casi con seguridad su imagen ha sido detectada en el Collado del Guijarral, un abrigo de Segura de la Sierra (Jaén), donde motivos esquemáticos cuentan con el añadido figurativo de siete ejemplares, aunque, sin razones claras se han interpretado como gatos domésticos [87]. (Fig. 7)
–Úrsidos. Hasta el momento no han aparecido huesos de oso en ninguno de los yacimientos cacereños de la Edad del Cobre, lo cual extraña, pues siendo una especie peligrosa y competidora del ser humano como el lobo, cualquier indicio hubiera bastado para suponer que alguna vez se le dio caza. Su presencia en los mismos territorios donde se desenvolvía el hombre así lo demandaba, pero hasta la fecha el único testimonio, aparte del grabado paleolítico de úrsido en la cueva de la Mina [88], es una pintura localizada en 1987 dentro de la cueva del Cancho Juracao de Navezuelas. Su hallazgo tuvo lugar en las mismas fechas que otro panel situado en el exterior de la cueva, pero solo el último llegó a publicarse [89], porque la divergencia de la primera con respecto a todo lo que conocíamos entonces de la pintura esquemática tradicional en nuestra región, imponía cierta mesura a la hora de pronunciarse acerca del encaje temporal de la misma, que, dicho sea de paso, aún avanzamos como hipótesis.
La pintura la compone un único motivo, aunque sospechamos que la mancha situada en la parte superior derecha hubiera podido formar parte del conjunto. Se ubican sobre un plano rocoso muy irregular, a menos de medio metro del suelo, en el fondo de la cueva, casi en total oscuridad, donde pasa totalmente desapercibida. Su propia morfología también se aparta de lo canónicamente establecido, pues lejos de la síntesis acostumbrada de las imágenes del Arte Esquemático, recorta la silueta del animal marcando desde la cabeza a la cola, la panza y las estilizadas patas. Su autor intentaba con ello ser lo más fiel posible a su naturaleza, aunque vacilante a la hora de concretar más detalles anatómicos. La percepción del perfil así, resalta con una corporeidad inédita, virando hacia un grado de naturalismo poco habitual en los álbumes del arte rupestre cacereño. Su color es rojo vinoso ligeramente desvaído, afortunadamente un tratamiento de la imagen, revela un intento de relleno o al menos un coloreado más denso sobre la cruz del animal, apreciándose, eso sí, una mayor fortaleza en el color del trazo que recorre la silueta, siendo esta una línea regular y gruesa. Los rellenos corporales con tintas planas son muy poco frecuentes, y como tal ha sido destacado recientemente como un rasgo sino insólito nada usual en el arte esquemático típico [90].
Muy interesante la mancha de color rojo intenso que parece salir de su boca, pues son muy raras las pinturas de esta zona que incorporen detalles complementarios de la acción que se pretende reflejar, apreciándose con ello cierta intención narrativa, tal vez, la representación de un animal herido. Hoy sabemos que figuras naturalistas y esquemáticas llegaron a coexistir [91], pero el carácter único de esta figura en su espacio territorial, complica un encuadre temporal llamado a recular en el tiempo y manteniéndonos a la expectativa, a la hora de explicar estos extraños desajustes de estilo y contexto. No cabría en este caso remontarse al sustrato iconográfico epipaleolítico, pues, aunque tiene una base reconocida en la región, los criterios que lo definen [92], solo se ve reflejado en una ínfima parte, tendríamos por tanto que considerar como posible, una realización posterior, tal vez en los momentos previos a la eclosión de las figuras propiamente esquemáticas, o hasta poder sumar más pruebas, especular incluso con el derecho a su excentricidad, mientras tanto cabría asignarle el calificativo de seminaturalista. (Fig. 8)
La imagen del oso no es inédita totalmente en la pintura esquemática extremeña, al menos, si se tiene en cuenta la idoneidad de su representación en una pintura de la sierra de Magacela [93]. No obstante, en la pintura paleolítica y en el arte levantino su representación era igualmente escasa y curiosamente coincidentes con las primeras en su ubicación en fondos de gruta [94]. Desde una perspectiva antropológica y mitológica, su figura ha sido tratada desde diferentes perspectivas resaltando sus dones como animal provisor de la sabiduría, demiurgo, de transcendental importancia en algunas ceremonias y relatos de los pueblos primitivos, donde la occisión o muerte violenta del animal tiene por misión garantizar la salud y la abundancia en el grupo humano [95].
–Otros. Podríamos apurar un poco más en contenido de algunas pinturas, pero, aunque estemos hasta cierto punto seguros que se dibujaron peces y lepóridos en el Paso de Pablo y en el Risquillo de Paulino, la proporción con la que también trabajan los autores de las pinturas, rebaja aún más las posibilidades de un reconocimiento fidedigno. Idéntico rasero puede aplicarse a los motivos cruciformes del abrigo de los Vencejos, pues podrían abstraer la imagen de un ave. Con los ofidios ocurre algo parecido, pues con cierta reticencia se les ha querido ver retratados en las líneas sinuosas de la cueva de los Cabritos de Berzocana y en panel nº 17 de Navaluenga en Peraleda de San Román [96] mientras que, al formar parte del conjunto de grafías utilizadas como recurso frecuente en la decoración de los megalitos, no ofrece la menor duda el diseño zizagueante en un ortostato de la Coraja.
5. CONSIDERACIONES FINALES.
Teniendo presente todos estos datos, podemos proporcionar en apretada síntesis, un resumen de distintas perspectivas que tenemos sobre el comportamiento humano en relación a la fauna de un periodo comprendido entre la aparición en nuestra región de las primeras comunidades productoras de alimentos y su consolidación como sociedades con un nivel de complejidad. La primera consecuencia es la fragmentación del ecosistema entre lo que podríamos denominar un espacio domesticado y otro asilvestrado donde los animales asumen un protagonismo distinto.
El espacio domesticado funciona con arreglo a un régimen económico de estructura mixta, basado en primer lugar, en un sistema de producción agroganadera, apegado aún a las viejas prácticas recolectoras estacionales de caza y pesca, evidentemente complementadas con actividades agrícolas, artesanales y de comercio cuyo desarrollo permitirían reconocer el nivel de desarrollo tecnológico alcanzado. En este sistema los componentes materiales y el control de espacios de óptima calidad, parece dirigido hacia la ganadería, que priorizaría por necesidades adaptativas la cabra y/o la oveja sobre el ganado porcino, caballar u bovino, desde época muy temprana.
Estas especies van a estar presentes desde el Neolítico en la región y variarán el régimen de apetencia de los habitantes de los poblados en función de las características del medio, que a su vez es transformado de una forma armónica para darles cabida. Durante la fase plena de la Edad del Cobre, estas prácticas están consolidadas y se observan escasas variaciones de unos con respecto a otros que pueden ser el origen de especializaciones locales.
La actividad cinegética desde el Neolítico a la Edad del Bronce sufre un descenso paulatino, a medida que crece el grado de complejidad del sistema económico. La persistencia sin embargo de especies cinegéticas en todos los niveles, aconseja interpretarlas como complemento dietético, aunque por las razones ya aducidas, tampoco puede separarse de las prácticas rituales, tal vez como una prueba de las habilidades del cazador, cuyo testimonio aparece implícito en las representaciones del arte rupestre o de operaciones de descaste, impelidas por la necesidad de evitar daños a los cultivos y al ganado, o la simple eliminación de competidores.
En las parrillas de los yacimientos, los porcentajes de especies salvajes contrastan considerablemente. El ciervo es el animal preferido por antonomasia, superado en tan solo una ocasión por los lepóridos en el yacimiento de los Barruecos, lo cual habla también de las intensas transformaciones que estaba sufriendo el ecosistema. Los índices menores del resto de las especies son asimismo el resultado de una caza ocasional de descaste o con intenciones de aprovechamientos menores.
En el dominio silvestre, los testimonios pictóricos son mucho más asimétricos, pues, por una parte, tenemos como el discurso pictórico se hace eco de lo cotidiano a través de escenas cinegéticas y de pastoreo con presencia destacada de cérvidos y cápridos, con cuya representación, se enfatiza la creciente capacidad de los grupos humanos de intervenir sobre la naturaleza utilizando los símbolos de la domesticación y la caza. Este protagonismo se ve respaldado a su vez por los datos de los asentamientos, pues ambos animales en sus respectivos lados de la tabla, lideran los índices de consumo porcentual.
El resto de los animales, sin embargo, tiene una representación mucho menos numerosa pero igualmente amplia con lo que respecta al número de especies, y como no podía ser de otra manera, se harán presentes las que tienen un mayor encaje ambiental. No es de extrañar entonces que cabras montesas, jabalí, gato montés, lobos, zorros u osos, tengan una oportunidad similar de comparecer en los paños rocosos de las serratas cuarcíticas.
El papel de los perros, acoplados al ámbito civilizado, harán su aparición en contextos de caza o pastoreo justificadas por la presencia humana, y la vaca o el caballo, como representantes del espacio domesticado, prestarán allí su imagen de una forma casi testimonial.
La versión estadística dice que se pintan sobre todo especies que se consumen, pero en el nicho ecológico mucho más diverso de las serranías y de los batolitos graníticos, donde se concentran los recursos complementarios para la subsistencia fuera del territorio habitualmente explotado, es inevitable que entren en juego otros detalles para hacer más comprensible su cosmovisión. Allí, el componente ideológico sale del marco de la supervivencia, sirviéndose de relatos pintados como una forma de legitimar el control ejercido sobre esos territorios. Paradójicamente, la adopción de formas de explotación agropecuaria, vino a reforzar el papel simbólico de las especies de fauna salvaje, donde su caza iba más allá de del mero acopio alimentario para convertirse en un acto de prestigio y símbolo de unidad grupal, con toda una serie de cualidades metafóricas que emanan de esa actividad. Recíprocamente y como contrapunto, las escenas donde se reflejan claramente actividades ganaderas, marcarían el límite de lo que fue una economía exclusivamente depredadora y sublimarían el logro de las nuevas metas civilizadoras, donde no olvidemos los antropomorfos, como expresión de la condición humana, son ahora el centro de ese universo.
El bestiario presente en el repertorio iconográfico de las pinturas cacereñas puede por tanto ser muy limitado en él número, pero en lo temático, es mucho más diversificado de lo que cabría esperar, pues no se limita a la plasmación de las figuras económicamente interesantes para al hombre, sino a la propia naturaleza contemporánea del mismo, estas instantáneas refuerzan su valor, cuando las contrastamos con el dato arqueológico preciso de la fauna documentada en los asentamientos, ofreciendo la posibilidad de un contrapunto a las cronologías ya de por si relativas del Arte Esquemático.
NOTAS.
[1] Cerrillo, C., E. et al. 2006: Los Barruecos: primeros resultados sobre el poblamiento neolítico de la cuenca extremeña del Tajo, Memorias de Arqueología Extremeña 6. Mérida.
[2] Enríquez Navascués, J. J. y García Cabezas M. 2011: Excavaciones arqueológicas en los yacimientos calcolíticos de Torrequemada y Torreorgaz (Cáceres). From the Origins: The Prehistory of the Inner Tagus Region. BAR, 2219. Oxford. Pp. 219-232.
[3] Castaños Ugarte, P. 1991: Animales domésticos y salvajes en Extremadura. Origen y evolución. Revista de Estudios Extremeños XLVII, 1. Badajoz. Pp. 9-66; Castaños Ugarte, P. 1992: Estudio Arqueozoológico de la fauna del Cerro de la Horca (Plasenzuela, Cáceres). Archeofauna 1. Pp.127-146. González Cordero, A. 2011: La Edad del Cobre en la Alta Extremadura. Asentamientos y organización del territorio. Facultad de Filosofía y Letras de la Universidad de Extremadura. Tesis Doctoral. Vol. I y II. Cáceres.
[4] Otros análisis de fauna se han llevado a cabo sobre otro tipo de yacimientos, siendo un ejemplo, por el nivel de profundidad alcanzado, una serie de aproximaciones zooarqueológicas a la fauna pleistocénica de Maltravieso (Canals, A.; Van der Mader, J.; Sauceda, Mª. I. y Carbonell, E. 2003: El conjunto paleontológico de la cueva de Maltravieso (Cáceres). XI Reunión Nacional de Cuaternario. Oviedo. Pp. 313-320; Rodríguez Hidalgo, A. J.; Muñoz Encinar, L. y Canals i Salomó A. 2008: Una aproximación zooarqueológica al yacimiento de la cueva de Maltravieso. El Mensaje de Maltravieso 50 años después (1956-2006). Memorias del Museo de Cáceres, 8. Cáceres. Pp. 153-163; Muñoz et al., 2008. Op. cit y más recientemente la de la cueva de Santa Ana.
[5] Castaños, P. 1991.Op. cit.
[6] Cerrillo Cuenca, E. y González Cordero, A. 2007: Cuevas para la eternidad: sepulcros prehistóricos de la provincia de Cáceres. Ataecina, 3. Instituto de Arqueología de Mérida.
[7] El yacimiento de Los Barruecos dispone de dos análisis faunísticos, en el más reciente de Arturo Morales prestó a atención a la fauna recogida en los niveles Neolíticos (Morales Muñiz, A. 2006: Los Barruecos (Malpartida de Cáceres): una fauna neolítica a cielo abierto. Los Barruecos: primeros resultados sobre el poblamiento Neolítico de la Cuenca Extremeña del Tajo. Memorias de Arqueología Extremeña, 6. Mérida. Pp. 233-420 y el más antiguo de Pedro Castaños, lo dedicó a la fauna extraída en niveles correspondientes a la Edad del Cobre Castaños, 1991. Op. cit. Pp. 20-22.
[8] González, A. 2011. Op. cit, p. 556.
[9] Fortea Pérez, J. y Martí Oliver, B. 1984: Consideraciones sobre los inicios del Neolítico en el Mediterráneo español. Zephyrvs XXXVII-XXXVIII. Salamanca. P. 188.
[10] Olària Pujoles, C. 2008-2009: Las mujeres y los orígenes de la domesticación. Primeros testimonios de la transformación económica en el arco mediterráneo peninsular del 11000 BP al 7000 BP. Cuadernos de Arte Rupestre, 5. Murcia. P. 46.
[11] Rubio, I. 1988: La economía de subsistencia en el Neolítico hispano, (P. López Coord), El Neolítico en España. Madrid. Pp. 379.
[12] Gavilán Ceballos, B. 1991: Análisis macroespacial de ocho yacimientos neolíticos en cueva de la subbética cordobesa: una contribución al estudio de la explotación de recursos durante la prehistoria. Cuadernos de Prehistoria de la Universidad de Granada 18. Madrid. P. 48.
[13] Asquerino Fernández, Mª. D. 1983: Una aproximación a la paleoeconomía del Neolítico: La cueva del Nacimiento (Pontones, Jaén). Homenaje a Martín Almagro. Madrid. P. 123.
[14] Morales, A. 2006. Op. cit. P. 129.
[15] Castaños, P. 1992.Op. cit.
[16] Martín Morales, C. 1987: El poblado de Almizaraque. Los inicios de la Metalurgia. El origen de la metalurgia en la Península Ibérica I. Oviedo. P. 15.
[17] Harrison, R. J. 1984: Beaker cultures of Iberia, France and the west Mediterranean island. L´Age du cuivre civilisations a vases campaniformes. París. P. 134.
[18] Martín de la Cruz, J. C. 1994: Avance en la discusión sobre medio ambiente en La Prehistoria. La relación vegetación fauna en el caso de Papauvas (Aljaraque, Huelva). Arqueología en el entorno del Bajo Guadiana.Huelva.P. 203.
[19] Chapman, R. 1991: La formación de las sociedades complejas. Barcelona. P. 187.
[20]Morales, A.2006. Op. cit. P. 120.
[21] Rodríguez y García, 2011. Op. cit. P. 235.
[22] Acosta, P. 1963: Estado actual de la prehistoria andaluza: Neolítico y Calcolítico. Habis,14. Sevilla. Pp. 195-105.
[23] Moreno Garçía, M. 2010: Estudo Arqueozoológico dos restos faunísticos do povoado Calcolítico do Mercador (Mourão). As comunidades agropastoris na margen esquerda do Guadiana. 2ª metado do IV aos inícios do II milenio AC. (Edic A. Valera). Memorias d´Odiana. 2ª Série. P. 324.
[24] Castaños, P. 1991. Op. cit. P. 138 y Rodríguez y García, 2011. Op. cit. P. 236.
[25] Castaños Ugarte, P. 1997: Estudio de la fauna del Cerro I de Los Castillejos (Fuente de Cantos, Badajoz). Norba. Revista de Historia, 14. Cáceres. P. 46.
[26] Castaños, 1998. Op. cit. P. 65.
[27] Pérez Ripoll M. 1999: La explotación ganadera durante el III milenio a. C. en la Península Ibérica. Sagvntum, Extra 2, Valencia. P. 97.
[28] Harrison, R. J. y Moreno, G. 1985: El policultivo ganadero o la revolución de los productos secundarios. Trabajos de Prehistoria, 42. Madrid. P. 64.
[29] Almagro Gorbea, M. y Martín Bravo, A. Mª. 1994: Medellín 1991, la ladera norte del Cerro del Castillo. Castros y oppida en Extremadura. Complutum Extra 4. Madrid. P. 121.
[30] Driesch, V. de A. y Boessneck, J. (1976): Die fauna von Castro do Zambujal. Studien über früe Tierknochenfunde von Iberischen Halbinsel 5. Mainz. Pp. 4-129; Schüle, W. (1969): Archaologie und Biologie. Forschungsberichte 15. Glockenbercher und Hauspferde en J. Boesssneck y Harrison, R. J. 1984. Op. cit. P. 136.
[31] Rodríguez y García, 2011. Op. cit. P. 235.
[32] Pellicer, M. y Acosta, P. 1982: El Neolítico Antiguo en Andalucía Occidental. Actes du Colloque International de Prehistoire (Montpellier, 1981): Archeologie en Languedoc. Montpellier. Pp. 49-60.
[33] Arribas, A. y Molina, F. 1979: El poblado de Los Castillejos en las Peñas de los Gitanos (Montefrío, Granada), Campaña de excavaciones de 1971, El corte, núm. 1. Cuadernos de Prehistoria de la Universidad de Granada, Serie Monográfica, 3. Granada. Pp. 152-168.
[34] Uerpmann, H. P. 1995: Domestication of the horse -when, where, and why?. Le cheval et les autres équidés: aspects de l’histoire de leur insertion dans les activités humaines, Colloques d’Histoire des Connaissances Zoologiques (Université de Liège). Pp. 15-29; Cardoso, J. L. y Detry, C. 2001-02: Estudo zooarqueologico dos restos de ungulados do povoado pre-historico de Leceia (Oeiras). Estudos Arqueologicos de Oeiras 10. 131-182.
[35] Pérez Ripoll, M. 1987: Evolución de la fauna prehistórica en el Mediterráneo español: Metodología, técnicas de troceado y su interpretación arqueológica. Tesis Doctoral. Universidad de Valencia inédita.
[36] Castaños, P. 1992. Op. cit. P. 133.
[37] Castaños, P. 1991. Op. cit. P. 34.
[38] Driesch, V. de A; Boessneck, J.; Kokabi, H y Schaeffer H. 1985: Osteologische Besonderheiten von Morro de la Mezquitilla (Málaga). Madrider Mitteilungen, 26. Mainz. Pp. 23-65; Altuna, J. 1980: Historia de la domesticación en el País Vasco desde sus orígenes hasta la Romanización. Munibe 32. Vitoria. Pp. 1-163 y Castaño, 1992. Op. cit.
[39] Valera, A.; Nunes, T. y Costa, C. 2010: Enterramientos de canídeos no neolítico: a fossa 5 de Corça 1 (Brinches, Serpa). Apontamientos de Arqueología e Patrimonio, 5. Pp. 12-15.
[40] Lomba Maurandi, J., López Martínez, M., Ramos Martínez, F. y Avilés Fernández, A. (2009): El enterramiento múltiple, calcolítico, de Camino del Molino (Caravaca, Murcia). Metodología y primeros resultados de un yacimiento excepcional. Trabajos de Prehistoria. Vol. 66 (2), Madrid. Pp.143- 159.
[41] Sanchís A. y Sarrión, I. 2004: Restos de cánidos (Canis familiaris ssp.) en yacimientos valencianos de la Edad de Bronce, Archivo de Prehistoria Levantina Vol. XXV, Valencia. Pp.161-198.
[42] Castaños, P. 1992. Op. cit. P. 141.
[43] Asquerino, Mª. D. y López, P. 1981: La cueva del Nacimiento (Pontones, yacimiento Neolítico en la Sierra de Segura. Trabajos de Prehistoria, 38. Madrid. P. 144.
[44] Driesch et al., 1985. Op. cit. P. 43.
[45] González Cordero, A. y Cerrillo Cuenca, E. 2016: Pesca y recolección de molusco en el Campo Arañuelo y Los Ibores durante la prehistoria reciente. XXII Coloquios Histórico Culturales del Campo Arañuelo. Navalmoral de la Mata. Pp. 167-192.
[46] González Cordero, A. y Cerrillo Cuenca, E. 2015: Los anzuelos de la cueva de la Canaleja (Romangordo (Cáceres). Arpi, 3, Homenaje a Rodrigo de Balbín Behrmann. Alcalá de Henares. Pp. 56-72.
[47] Rodríguez y García, 2011. Op. cit. P. 236.
[48] Martí Oliver, B; Arias Gago, A; Martínez, R. y Cabanilles, J. 2001: Los tubos de hueso de la Cova de L´Or en Beniarrés, Alicante). Instrumentos musicales en el Neolítico Antiguo de la Península Ibérica. Trabajos de Prehistoria, 58, 2. Madrid. P. 41.
[49] Castaños, P. 1992. Op.cit. 130.
[50] Tavares da Silva, C. 1987: Megalitismo do Alentejo occidental e do sul baixo Alentejo (Portugal). El megalitismo en la Peninsula Ibérica. Madrid. Pp. 85-95.
[51] Gusi y Jener, F. y Olària y Pujoles, C. 1991: El poblado neoeneolítico de Terrera Ventura (Tabernas, Almería). Excavaciones Arqueológicas de España 160. Madrid. P. 22.
[52] Pavón Soldevila, I. 1998: El Cerro de Alange (Badajoz). Intervenciones arqueológicas. Memorias de Arqueología Extremeña 1. Mérida. P. 167.
[53] Arribas y Molina, 1979. Op. cit. P. 157.
[54] Tavares, C. 1987. Op.cit. P. 80.
[55] García Arranz, J. J. 1990a: Los animales en la pintura rupestre esquemática de las Villuercas (Cáceres): testimonio gráfico de un modo de vida prehistórico. Alcántara (tercera época), 20. Cáceres. Pp. 89-110.
[56] García, 1990ª. Op. cit. P. 93.
[57] Rivero de la Higuera, Mª. C. 1972-73: Nuevas estaciones de pintura rupestre esquemática en Extremadura. Zephyrvs, XXIII-XXIV. Salamanca. P. 305, Fig. 10.
[58] García, 1990ª. Op. cit. P. 97.
[59] Grande del Brío, R. 1980: Nuevos abrigos con pinturas rupestres en Monfragüe (Sierra de las Corchuelas, Cáceres). Zephyrvs, XXX-XXXI. Salamanca. Pp. 147-152.
[60] Alvarado Gonzalo, M. De y González Cordero, A. 1979: Pinturas esquemáticas en Malpartida de Cáceres. Revista Alcántara. 1ª época, nº 195, año XXXV. Cáceres. Pp. 16-22.
[61] Pastor González, V. y Pastor Novella, Mª. T. 2014: XLIII Coloquios Históricos de Extremadura. Trujillo. En:https://www.chdetrujillo.com/wp-content/uploads/2014/Pastor-Gonzalez-Vicente-e-hija.pdf.
[62] Collado Giraldo, H. y García Arranz, J. J. (2007): últimas investigaciones en la cueva del Castillo de Monfragüe (Cáceres). Actuaciones de adecuación para las visitas y revisión de sus manifestaciones rupestres. Cuadernos de Arte Rupestre, 4. Murcia. Pp. 313-351.
[63] Pastor V. y Pastor Mª. T., 2014. Op. cit. P. 29. Fig. 26
[64] González Cordero, A. y Alvarado Gonzalo, M. de 1993: Nuevas pinturas rupestres en Extremadura. Pintura naturalista en el entramado esquemático de las Villuercas (Cáceres). Revista de Arqueología. 143. Madrid. Pp. 18-25.
[65] Olària, C., 2008-2009. Op. cit.
[66] Jordá Cerdá, F. 1975: La Peña del Escrito (Villar del Humo, Cuenca) y el culto al toro. Cuadernos de Prehistoria y Arqueología Castellonenses, Número 2. Castellón de la Plana. P. 9.
[67] Piñón Varela, F. 1982: Las pinturas rupestres de Albarracín (Teruel), Centro de Investigaciones y Museo de Altamira. Monografías núm. 6. Santander. P. 207.
[68] Beltrán Martínez A. 1968: Sobre la pintura rupestre levantina de un caballo cazado a lazo del abrigo de Selva Pascuala en Villar del Humo (Cuenca). Miscelánea Lacarra, Zaragoza, P. 81.
[69] Ruiz López, J. F. 2009: Cazadores y Presas: simbolismo e interpretación social de las actividades cinegéticas en el arte levantino. Arqueobios. 3. Vol I. diciembre. P. 111.
[70] Breuil, H. 1933: Les peintures rupestres schématiques de la Péninsule Ibérique. Vol. III,
Sierra Morena, Lagny.
[71] López Payer, M. y Soria Lerma, M. 1991: Análisis estilístico de los conjuntos del Canjorro de Peñarrubia, Doña Clotilde, Abrigo del Tío Campano y Selva Pascuala. Espacio, Tiempo y Forma. Serie I, Prehistoria y Arqueología, Vol. IV. Madrid. P. 235.
[72] García Arranz, J. J. 2015: El arte rupestre en el Geoparque Villuercas Ibores Jara (Cáceres). Dossier Curso Guía Ranger del Geoparque Villuercas Ibores Jara, mayo 2015. P. 4.
https://www.academia.edu/12346909/El_arte_rupestre_en_el_Geoparque_Villuercas
[73] García, J. J. 1990a. Op. cit. P.105, Fig. X.
[74] García Arranz, J. J. (1990b): La pintura rupestre esquemática en la comarca de las Villuercas (Cáceres). Edic. Institución Cultural el Brocense. Cáceres. P. 123.
[75] Ripoll López, S. y Collado Giraldo, H. 1996: Una nueva estación paleolítica en Extremadura. Los grabados de la cueva de la Mina de Ibor (Castañar de Ibor (Cáceres. Revista de Estudios Extremeños, LII, 2. Badajoz. Pp. 383-399.
[76] González, A. y De Alvarado, M. 1993. Op. cit.
[77] Rivero, Mª. C. 1972-73. Op. cit. P. 289.
[78] Vázquez Varela, J. M. 1990: Petroglifos de Galicia. Biblioteca divulgativa, Serie Galicia, 3. Santiago de Compostela. P. 104.
[79] García, J. J. 1990ª. Op.cit. P. 106 Fig. XI.
[80] Ruiz, J. F. 2009. Op. cit. P. 114.
[81] Pastor, V. y Pastor, Mª. T. 2014. Op. cit. P. 24 y 29.
[82] González Cordero, A. y Quijada González, D. (1991): Los Orígenes del Campo Arañuelo y la Jara cacereña. Su integración en la prehistoria regional. Navalmoral de la Mata. P. 136.
[83] Jiménez Lorente, S. y Ayala Juan, Mª. M. 2006: Avance a la representación del canis familiaris en la pintura rupestre postpaleolítica. Cuadernos de Arte Rupestre, 3. Murcia. Pp. 161-184.
[84] González Cordero, A. y de Alvarado Gonzalo, M. 1997: Nuevos conjuntos esquemáticos de las Villuercas cacereñas. II Congreso de Arqueología Peninsular. Vol. II. Zamora. P. 25.
[85] García Arranz J. J.; Collado, H.; Fernández, M.; Girón, M.; García, Mª I. y Mesa, Mª. J. 2011: La cueva Chiquita o de Álvarez (Cañamero, Cáceres): recientes intervenciones y revisión de sus manifestaciones rupestres. Espacio, Tiempo y Forma. Serie I. Nueva época. Prehistoria y Arqueología, Tomo 4. Madrid. P. 91
[86] Las pinturas de la cueva de los Horcones fueron descubiertas por Jaime Cerezo y José Pedro Cortijo. Al primero debemos además las fotos en gran detalle de este lugar por lo que, en prueba de agradecimiento, queremos dejar constancia en estas páginas
[87] Ayala Juan, Mª. M. y Jiménez Lorente, S. (2005): Las pinturas rupestres del Collado del Guijarral (Segura de la Sierra, Jaén). Cuadernos de Arte Rupestre, 2. Murcia. Pp. 212.
[88] Ripoll, S. y Collado, H. 1996. Op. cit. P. 386.
[89] González, A. y de Alvarado, M. 1997. Op. cit. P. 289.
[90] Collado Giraldo H. y García Arranz, J. J. 2010: Reflexiones sobre la fase inicial de arte rupestre esquemático en Extremadura a raíz de las recientes investigaciones. Actas del II Congreso de Arte Rupestre Esquemático en la Península Ibérica. Almería. P. 293.
[91] Mas Cornellà, M. 2001: Estructuras iconográficas e identificación de especies (secuencias iniciales y finales del arte postpaleolítico “esquemático). Quaderns de Prehistòria i Arqueologia de Castelló, 22. P. 169.
[92] Collado Giraldo, H. y García Arranz, J. J. 2012: La revalorización del arte rupestre de los grupos depredadores postpaleoliticos en la Península Ibérica: el arte rupestre preesquemático. El problema “Levantino” Arte rupestre postpaleolítico en la Península. Ibérica. Budapest. P. 258.
[93] Collado Giraldo. H. 1995: Sistematización cronológica de la pintura rupestre esquemática en la provincia de Badajoz: los abrigos de la Sierra de Magacela. Espacio Tiempo y Forma, Serie I, Prehistoria y Arqueología, t. 8. Madrid. P. 154-grupo A. VIII.2. Fig. 12.
[94] Leroi-Gourhan, A. 1972: Considerations sur l´organization spatiales des figures animales dans l’art parietal paleolithique. Actas del Symposium Internacional de Arte Prehistórico. Santander. P. 283.
[95] Jordán, Montes, J. F. y Molina Gómez, J. A. 2007: Los osos en el arte rupestre postpaleolítico español. ¿Un mito de la resurrección y de la fertilidad? Cuadernos de Arte Rupestre, 4. Murcia. P. 233.
[96] González Cordero, A. (2009): De los paisajes sagrados a los espacios simbólicos. El santuario rupestre del valle de Cancho Castillo (Peraleda de San Román). XV Coloquios Histórico Culturales del Campo Arañuelo. Navalmoral de la Mata. Fig. 1.
ÍNDICE DE FIGURAS

1-Representaciones de cápridos en el Cancho de la Burra en Cañamero (Dcha.) y el Paso de Pablo en Roturas (Izqda.). Fot. Antonio González.

2- Cuartos traseros de un équido representados en el abrigo Florencio (Retamosa). Fot. Filtrada a través de DStretch por V. Pastor y Mª. T. Pastor (2014).

3-Figura femenina sujetando un bóvido con una cuerda. Panel del Paso de Pablo (Roturas). Fot. Antonio González.

4-Cervidos representados en el abrigo de la Madrastra 1 de Cañamero (Dcha.) y Cérvido del Paso de Pablo en Roturas (Izqda). Imagen filtrada para resaltar las figuras en negro. . Fot. Antonio González y E. Cerrillo C.

5-Dos ejemplares de Capra hispánica representados en el abrigo Florencio de Retamosa, Filtrada con DStretch por V. Pastor y Mª. T. Pastor (2014).

6-Tres cánidos representados en la parte inferior del panel de Paso de Pablo (Roturas) junto a un cáprido. Fot. Antonio González.

7-Imagen de un gato montés de la cueva de Los Horcones en Roturas. Fot. de Jaime Cerezo.

8-Imagen de un úrsido pintado en el interior de la cueva del Cancho Juracao de Roturas. Filtro DStretch y tratamiento posterior de imagen. Fot. Antonio González.

